К.Тринчер. О воде и теплокровности.
Опубликовано в SANUM-Post № 15/1991, стр. 21-26.
Перевод с немецкого Н. Луковниковой под ред. Р. Минвалеева
О воде и теплокровности
Вода все еще остается тайной жизни
Проф., доктор медицины Карл Тринчер
Вода представляет собой растворитель, в котором, как бы странно это ни звучало, растворяются все вещества, и даже золото растворимо в воде. Чтобы это понять, необходимо, по выражению великого химика Фрица Хабера, иметь правильное химическое чутье. После окончания Первой Мировой войны в морях появилось поразительно много небольших торговых кораблей под немецкими флагами. Они брали пробы воды, которые затем проверялись в лабораториях на наличие золота. Содержание золота оказалось достаточно велико, чтобы техническим способом начать добычу золота из морской воды. Это стало бы для побежденных немцев выгодным предприятием, поскольку тогда они смогли бы выплачивать наложенные репарации золотом. Послевоенная политика решила все иначе. Репарационные платежи, наложенные на немцев, которые они могли бы выплачивать технически добытым золотом, были отменены, поэтому дальнейшие поиски золота в морской воде были прекращены. Но научный результат был достигнут, а именно, было доказано, что драгоценный металл – золото - растворимо в воде.
Чужеродные молекулы меняют плотность воды
Однако, если наличие золота в воде доказано, то возникает вопрос, действительно ли все вещества растворимы в воде. Тогда с водой было проведено множество экспериментов, и вдруг, как гром среди ясного неба, в научных публикациях появился новый термин «вода Дерягина». Плотность вещества относится к его главной характеристике. Также хорошо известна плотность воды и ее зависимость от температуры и давления. Теперь же открытие русского физика Дерягина потрясло научный мир. Он доказал, что плотность воды в очень узкой капиллярной трубке из кварца выше, чем нормальное значение. Это выглядело так, будто в тесноте капилляра вода сжималась.
На протяжении двух лет обсуждения «воды Дерягина» в научной литературе одновременно во многих лабораториях повторяли опыт Дерягина и всегда с тем же исходом. Но, несмотря на более высокую плотность воды в узком капилляре, теоретики отказывались признавать этот результат. «Ваша вода с примесями», кричали они с негодованием, которое для меня было в новинку на научных конференциях. Снова и снова можно было услышать: «Вода у вас с примесями, и потому при замерах ее плотность выше». И они оказались правы, эти теоретики. Два года спустя один американский химик доказал, что «вода Дерягина» содержит частицы кварца. Содержащаяся в кварцевом капилляре вода растворяет в себе молекулы кварца стенок капилляра, становясь уже не чистой водой, а сильноразбавленным водным раствором кварца. Это естественным образом обусловливает более высокую плотность раствора, нежели плотность чистой воды.
Чистой воды не бывает
Чистая вода, состоящая только лишь из молекул Н2О, не существует; она всегда содержит растворенные вещества, а также она содержит частицы, из которых сама состоит. Это электрически заряженные частицы молекул воды: положительно заряженные ионы воды Н+ и отрицательно заряженные гидрокосильные ионы (ОН)–. В килограмме воды при температуре 22ºС содержится ровно 10–7 грамм Н+ и одновременно также 10–7 грамм (ОН)–. Обозначим величину 10–7 грамм ионов Н+ через символ [Н+]. Молекулярный вес воды, соответствующий одному молю воды, составляет18 грамм. Килограмм воды, таким образом, содержит:
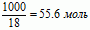
Один моль означает количество молекул, соответствующих молекулярному весу вещества. Обозначенное через N0 количество молекул в одном моле, так называемое число Авогадро, составляет величину N0 = 6.06·1023.
Если мы умножим содержащиеся в килограмме воды 10–7 грамм ионов Н+ на N0, мы получим число ионов Н+ в одном моле воды:
[Н+] · N0 = 10–7 · 6.06·1023 = 6.06·1016.
С другой стороны один литр воды содержит:
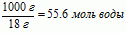
где 18 грамм – молекулярный вес воды.
Если мы умножим 55.6 на N0, то получим число молекул воды в одном литре воды:
Z(H2O) = 55.6 · 6.06·1023 = 336·1023.
Из отношения чисел [Н+] · N0 и Z(H2O) находим:
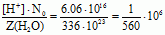
Это отношение показывает, таким образом, что только одна молекула воды на 560 миллионов молекул диссоциирует в соответствии с уравнением:
Н2О = Н+ + (ОН)–.
Таким образом, чистая вода в очень малой части диссоциирует на ионы. Данная величина точно определена и стала очень важной в химии и биологии. Продукт реакции, состоящий из ионов Н+ и (ОН)–, называется ионным произведением воды и выражается величиной КW, при 220С принимающей значение 1·1014. Данная величина изменяется при увеличении температуры, т.е. повышение температуры способствует диссоциации воды. Константа ионного произведения, формально выраженная, обозначается, таким образом:
КW = [Н+]·[ОН–] = 1·1014 (при 220С).
Из данного уравнения ясно, что есть три различных состояния воды в отношении содержания ионов:
Первое: Состояния нейтральности, когда наблюдается равенство концентрации ионов: [Н+] = [ОН–].
Второе: Состояние кислотности, представляющее собой неравенство концентрации ионов, а именно более высокую концентрацию ионов Н+: [Н+] > [ОН–].
Третье: Состояние щелочности, когда существует более высокая концентрация ионов (ОН–): [ОН–] > [Н+].
При нейтральной реакции для [Н+] будет действительна величина [Н+] = 10–7.
Для кислотной реакции действительно: [Н+] > 10–7.
Для щелочной реакции действительно: [Н+] < 10–7.
Роль концентрации ионов воды
Чтобы упростить запись выражения концентрации ионов воды, введен общепризнанный символ, который выведен из латинских слов «potentia» и «hydrogenium», символ рН. Он представляет собой отрицательный десятичный логарифм концентрации ионов воды: рН = – lgН+. Таким образом, для нейтральной реакции имеем значение рН: рН = – lg10–7 = 7, для кислотной реакции: рН < 7 и для щелочной реакции: рН > 7.
Теперь обратимся к вопросу, как ведут себя животные клетки в ответ на изменения внешней среды, которая характеризуется уровнем рН. Подходящим объектом исследования являются красные кровяные тельца млекопитающих (эритроциты), которые могут быть легко заимствованы из их естественной среды, плазмы крови, и помещены в искусственную среду, в изотонический раствор поваренной соли. В нем эритроцит живет еще ровно два часа, а именно до момента расхода имеющейся в эритроците глюкозы. Таким образом, предоставляется возможность проверить поведение эритроцита по отношению к изменению уровня рН в искусственной среде, которая изменялась исключительно путем добавления кислоты (HCl) или щелочи (NaOH) в физиологический раствор поваренной соли.
Результат этих исследований схематически представлен на рис.1.
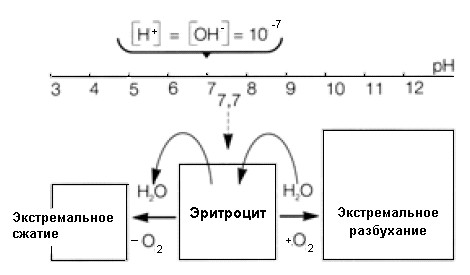
Рис.1 Зависимость изменения объема безъядерного эритроцита от рН среды.
Найденный в ходе экспериментов результат гласит: Эритроцит живет в слабощелочной среде с уровнем рН 7.7. При повышении щелочности среды эритроцит разбухает за счет накапления воды до экстремального разбухания при приблизительном уровне рН = 12. В среде с уровнем рН > 12 эритроцит лопается. При повышении кислотности среды эритроцит уменьшается в объеме за счет потери воды до своего минимального объема. Параллельно с процессом накопления воды эритроцитами происходит приток кислорода в слабощелочную среду респираторной легочной ткани. Параллельно с процессом потери воды в слабокислой среде, при контакте с клетками большого круга кровообращения, следует отдача кислорода. Таким образом, мы имеем здесь дело со своеобразным рН-обусловленным механизмом насоса эритроцита: приход кислорода связан с накоплением воды и протекает в щелочной среде легких, отдача кислорода связана с потерей воды и протекает в кислотной среде при контакте со слабокислыми клетками большого круга кровообращения.
Феномен теплокровности
Теперь обсудим второе значение щелочности респираторной легочной ткани, которое связано с энергетическим поведением теплокровного организма и сгоранием жиров в легких. С появлением теплокровных организмов была достигнута высшая ступень биологической эволюции. Температурный диапазон теплокровных ограничен и охватывает промежуток от 360С до 420С, причем млекопитающие расположились на нижней температурной границе, в то время как птицы с более интенсивным энергообменом подошли к верхней температурной границе теплокровности. Феномен теплокровности подводит нас к трем вопросам:
1. Почему состояние теплокровности является более высокой ступенью биологической эволюции в сравнении с состоянием хладнокровности?
2. Почему область температуры теплокровных ограничена промежутком от 360С до 420С?
3. Обладает ли организм специальным органом теплопродукции для поддержания состояния теплокровности?
1. На вопрос, почему появилось состояние теплокровности, можно ответить изречением Клода Бернара: «La fixité du milieu interieur est le condition de la vie libre». Физиологический смысл фразы «la vie libre» заключается в максимизации внутренней подвижности организма. В механистическом примере максимальная подвижность шара на поверхности заключается в максимальной подвижности шара на горизонтальной поверхности, на которой нет заданного направления движения. Постоянство температуры теплокровных организмов означает независимость протекающих в организме процессов от температуры внешней среды, хотя и связано с физиологическими процессами, которые обеспечивают постоянный уровень температуры. Независимость скорости биохимических процессов от температуры окружающей среды означает дополнительную степень свободы, которой не обладают хладнокровные организмы.
2. На вопрос, почему состояние теплокровности охватывает область от 360С до 420С, ответ следующий: температурная область состояния теплокровности обусловлена двумя экстремальными особенностями воды. Одна из этих особенностей – энергетическая – минимум удельной теплоемкости ср, вторая – механическая – максимум сжимаемости или деформируемости воды, которая характеризуется коэффициентом сжатия.
Зависимость удельной теплоемкости воды от температуры представлена на рис.2. В то время как для всех других веществ справедлива закономерность, что вещество должно тем больше принять тепла, чтобы его температура повысилась, чем выше его температура, для воды при температуре в промежутке от 00С до 450С имеет место обратная зависимость от температуры: чем выше температура, тем меньшее количество тепла необходимо для повышения температуры. В промежутке от 350С до 450С удельная теплоемкость воды принимает свое минимальное значение. Область минимального значения удельной теплоемкости, таким образом, является условием постоянства процессов теплокровного организма.
Зависимость сжимаемости воды от температуры можно увидеть на рис.3. В то время как для всех веществ, чем менее сжимаемо вещество, тем выше его температура, у воды при температуре от 00С до 450С обнаружена обратная закономерность: сжимаемость воды снижается при повышении температуры и достигает в температурном промежутке состояния теплокровности области минимальных значений. Вода, таким образом, в температурной области состояния теплокровности максимально деформируема, при этом деформируемость не зависит от температуры. Область максимальной деформируемости воды, характеризующаяся минимальным значением коэффициента сжатия на рис.3, является так же, как и область минимальных значений удельной теплоемкости (рис.2) условием постоянства процессов функционирования теплокровного организма.
3. На вопрос, есть ли у теплокровных организмов специализированный орган теплопродукции для поддержания функционирования теплокровного организма, ответ следующий: теплокровность обеспечивается специальным органом, легкими, которые помимо своей постоянной функции газообмена выполняют транзиторную температурнозависимую функцию теплопродукции.
Это явление легочного термогенеза теплокровных животных было экспериментально обнаружено еще в начале XX века, а именно, было доказано, что в легких окисляются жиры. Этот факт был зарегистрирован, но не проинтерпретирован. Лишь много позже усвоение жиров в легких было осознано как производство тепла (Тринчер, 1960).
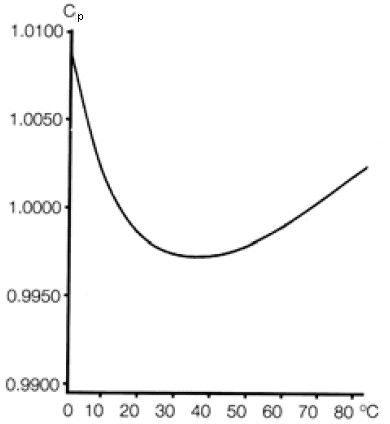
Рис.2 Зависимость удельной теплоемкости воды от температуры (Eisenberg & Kauzmann, 1969).
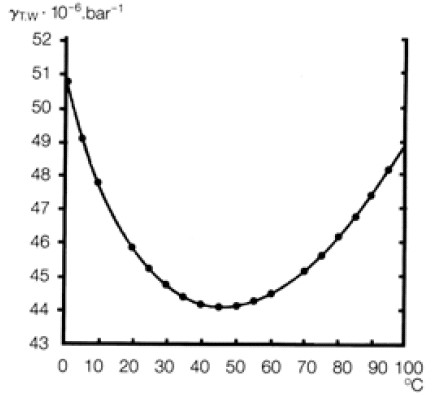
Рис. 3. Зависимость сжимаемости воды от температуры (Kell, 1967; Fine et al., 1973)
Значение «гипоксического парадокса»
Известно и доказано, что респираторная легочная ткань задерживает часть жировых капелек, эмульгированных в венозной крови, и что непосредственное окисление жиров кислородом вдыхаемого воздуха осуществляется в стенках респираторной легочной ткани.
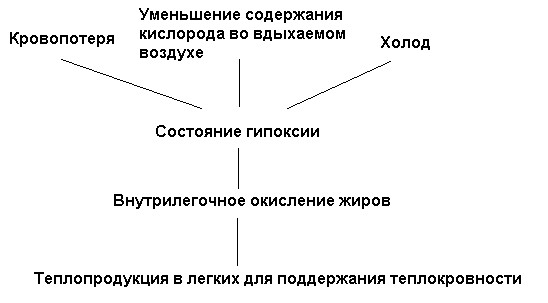
Такого рода опыты, доказывающие «усвоение жиров» в легких, были проведены в рамках почти необозримого количества физиологических и биохимических работ в различных вариантах. Особое значение, все же, имеет тот факт, что в состоянии гипоксии, т.е. при пониженном содержании кислорода во вдыхаемом воздухе, потребление кислорода теплокровным организмом возрастает. Это парадоксальное явление, заключающееся в том, что уменьшение содержания кислорода имеет своим следствием повышение потребления кислорода, было названо в соответствии со своей сутью «гипоксическим парадоксом». Экспериментально установлено, что состояние гипоксии также может быть вызвано острой кровопотерей. Теплокровный организм реагирует на острую кровопотерю физиологическим следствием в виде внутрилегочного окисления. К аналогичным физиологическим реакциям теплокровных организмов ведет состояние гипоксии. Таким образом, мы имеем следующие два идущие друг за другом состояния:
(А) Кровопотеря → состояние гипоксии → внутрилегочное окисление жиров.
(В) Снижение содержания кислорода во вдыхаемом воздухе → состояние гипоксии → внутрилегочное окисление жиров.
Последовательность (В) названа в физиологической литературе «гипоксическим парадоксом». Парадокс под этим названием заключается в следующем явлении: хотя вдыхаемый воздух содержит меньше кислорода, потребление кислорода теплокровного организма растет. Парадокс фактически состоит в том, что орган, получающий кислород из артериальной крови, реагирует на уменьшение поступления кислорода ростом его потребления.
Была высказана гипотеза, что умеренное снижение парциального давления кислорода выравнивается физиологическим возбуждением, которое побуждает клеточный метаболизм увеличить потребление кислорода. Неправдоподобность этой гипотезы только подчеркивает невозможность объяснить «гипоксический парадокс» в рамках известных физиологических законов. Таким образом, этот парадокс является своего рода призраком на эмпирическом поле физиологии. Для объяснения «гипоксического парадокса» требуются все же не гипотезы, но знание специфической способности легких реагировать на состояние гипоксии внутрилегочным окислением.
Важность теплопродукции легких
Адаптивным достижением теплокровных организмов, живущих в холодных условиях, является то, что они сохраняют квазипостоянство температуры тела путем выработки дополнительного тепла в респираторной ткани легких. При воздействии холода на теплокровные организмы для сохранения тепла теплокровные не имеют другой защиты кроме дополнительной теплопродукции путем сжигания жиров в легких. Огромная респираторная площадь легочной ткани является областью контакта между холодным альвеолярным воздухом и текущей по легочным капиллярам крови.
При воздействии экстремально сильного холода тепла, продуцируемого всеми дифференцированными клетками, а также дополнительного тепла, полученного за счет мускульной активности, недостаточно, чтобы предохранить текущую по легочным капиллярам теплую кровь от охлаждения холодным вдыхаемым воздухом. При таком экстремальном воздействии холода на теплокровный организм включается дополнительная теплопродукция в легких, связанная с окислением жиров в стенках альвеол. При этом высвободившееся тепло образует прослойку, некий теплый газовый фильтр, через который проходят в кровь холодные молекулы кислорода. Квазипостоянство температуры теплокровных организмов обеспечивается производством тепла в легких. Легкие создают тепловой барьер, закрывающий теплую кровь с постоянной температурой от воздействия холода вдыхаемого воздуха. В период эмбрионального развития теплокровные организмы находятся в естественном термостате, материнской утробе, и у них нет органа, функцией которого была бы выработка тепла, т.е. тепла как самоцели, но не как побочного продукта рабочих процессов. Легкие эмбриона не функционируют ни как орган дыхания, ни как орган теплопродукции. И только начиная с момента рождения, когда полностью завершается биологический переворот насильственного перехода от внутриутробной жизни к постнатальной, когда внезапно прекращается тепловая защита матки, новорожденному требуется новая тепловая защита, и тогда, в момент столкновения с холодным воздухом внешней среды, легкие начинают функционировать и как орган дыхания и, одновременно, как орган теплопродукции.
Действие холода на теплокровные организмы аналогично воздействию острой кровопотери и пониженного содержания кислорода во вдыхаемом воздухе. Холод вызывает периферическую гипоксию вследствие возрастания потребления кислорода в периферических регионах организма, так что поступающая в легочные капилляры кровь оказывается в состоянии холодовой гипоксии. Это гипоксическое состояние организма, вызванное воздействием холода, ведет к внутрилегочному окислению, сжиганию жиров в легких. Таким образом, наблюдается следующая последовательность:
(С) Воздействие холода → гипоксическое состояние → внутрилегочное окисление жиров.
Из представленных последовательностей (А), (В) и (С) вытекает непосредственная цель или физиологическая необходимость внутрилегочного окисления: оно служит сохранению гомойотермного состояния, состояния теплокровности. Теперь мы можем построить следующую схему, в которой представлена взаимосвязь между различными причинами гипоксии и внутрилегочным сжиганием жиров, служащим поддержанию теплокровности:
Резюме и выводы
1. Вода – универсальный растворитель; нет такого вещества, которое не было бы растворимо в воде.
2. Животные клетки живут в слабощелочной среде, в то время как сами клетки слабокислые. В щелочной среде клетки разбухают за счет накопления воды, в кислой среде клетки уменьшаются в размерах за счет потери воды.
3. Приток кислорода связан с накоплением воды и протекает в щелочной среде легких, отток кислорода связан с потерей воды и протекает при контакте со слабокислыми клетками большого круга кровообращения.
4. Температурный диапазон теплокровности от 360С до 420С обусловлен двумя экстремальными характеристиками воды: минимальной удельной теплоемкостью воды и максимальной сжимаемостью воды.
5. Легкие выполняют кроме своей постоянной функции газообмена также транзиторную дополнительную функцию теплопродукции.
6. Производство тепла в легких основывается на окислении жиров, когда теплокровный организм находится в состоянии гипоксии.
7. Состояние гипоксии теплокровных организмов может иметь различные причины: кровопотеря, снижение содержания кислорода в дыхательном воздухе, воздействие холода.
8. Так называемый гипоксический парадокс основывается на окислении жиров в легких в состоянии гипоксии.
Литература
1. Eisenberg, D., and Kauzmann, W. (1969): “The Structure and Properties of Water”,Oxford University Press.
2. Fine, R.A., and Millero, F.J. (1973): J. Chem. Phys. 59, 5529.
3. Kell, G.S. (1972): “Water and Aqueous Solutions”, Edition R.A. Horne, John Wiley and Sons,New York.
4. Trincher, K. (1981): “Die Gesetze der biologischen Thermodynamik”, Urban&Schwarzenberg, Wien; (1990): “Wasser - Grundstruktur des Lebens und Denkens”, Herder&Co., Wien.
Файлы для скачивания: